Van die bestaande lewende wesens is voëls en soogdiere homotermies (met die uitsondering van slegs blote molrotte). Daarbenewens is die eerste heeltemal warmbloedige vis op 15 Mei 2015 ontdek wat wetenskaplikes van die National Oceanic and Atmospheric Administration of the United States ontdek het. Die vraag of pterosaurusse en dinosourusse aan warmbloedige diere behoort het, kan ook bespreek word, hoewel navorsers onlangs meer en meer geneig is tot warmbloedigheid, en daar is reeds debatte oor watter van die spesies warmbloedig was en wat nie. Daar is ook geen finale duidelikheid oor watter soort endotermie die dinosourusse besit het nie, maar die beskikbare gegewens stel ons in staat om tot die gevolgtrekking te kom dat groot dinosourusse ten minste traagheidsgemeenskap gehad het.
Die meeste navorsers is van mening dat dinosourusse in hul metaboliese regime nie net 'n tussenposisie tussen "warmbloedige" en "koelbloedige" diere inneem nie, maar fundamenteel van albei verskil. Waarnemings van groot moderne reptiele het getoon dat as 'n dier 'n verminderde liggaamsgrootte van meer as 1 m het (naamlik, feitlik alle dinosourusse was so), dan is dit in 'n eweredige en warm (subtropiese) klimaat met klein daaglikse temperatuurskommelinge die vermoë om 'n konstante liggaamstemperatuur bo te hou 30 ° C: die hittekapasiteit van water (waarvan die liggaam 85% bestaan) is groot genoeg dat dit eenvoudig nie tyd het om oornag af te koel nie. Die belangrikste ding is dat hierdie hoë liggaamstemperatuur slegs verseker word vanweë hitte van buite, sonder enige betrokkenheid by hul eie metabolisme (waarvoor soogdiere 90% van die voedsel wat hulle verbruik, moet spandeer). Dus, 'n dier met groottes wat tipies is van die meeste dinosourusse, kan dieselfde mate van temperatuurbeheer as soogdiere bereik, terwyl dit 'n tipiese reptielmetaboliese tempo handhaaf, hierdie verskynsel J. Hotton (1980) wat traagheidshomeotermie genoem word. Blykbaar was dit die traagheidsgemeenskap (tesame met tweedrag) wat die dinosourusse tot die konings van die Mesosoïese aard gemaak het.
In 'n nuwe studie het Kanadese en Brasiliaanse wetenskaplikes moontlik 'n idee gekry vir hierdie evolusionêre raaisel. 'N Span gelei deur Glenn Tattersall van die Brock Universiteit het ontdek dat die Argentynse swart en wit tagu (Salvator merianae) het seisoenale warmbloedigheid. Hierdie akkedis, tot 150 sentimeter lank, woon in die grootste deel van Suid-Amerika en is goed bekend by bioloë. Soos die meeste ander reptiele bak die meeste van die jaar die tegue gedurende die dag in die son, en snags skuil hulle in gate en koel af. Wetenskaplikes wat sensors en hittekamers gebruik, het egter gevind dat gedurende die broeiseisoen, van September tot Desember, in die oggendure, die asemhalingstempo en hartklop van die dier toeneem, en hul temperatuur styg en met tien grade Celsius hoër as die temperatuur in die gat word. Wetenskaplikes glo dat Suid-Amerikaanse akkedisse 'n tussenskakel is tussen koelbloedige en warmbloedige diere. 'N Verhoging in liggaamstemperatuur gedurende die broeiseisoen verhoog hul aktiwiteit as u na 'n maat soek, versnel die ontwikkeling van eiers en laat u meer sorg vir die nageslag. Daarbenewens, byvoorbeeld, hou 'n leerskilpad, as gevolg van die werk van spiere, 'n isolerende vetlaag en groot groottes, 'n liggaamstemperatuur hoër as die temperatuur van die omringende water. Groot monitor akkedisse word ook warm tydens jag of aktiewe beweging. Groot slange soos pythons en boas kan die liggaamstemperatuur verhoog deur in 'n ring te krul en spiere saam te trek. Dit word gebruik om eiers op te warm en uit te broei.
Tipes homeotermie
onderskei waar en traagheid homeothermy.
- Ware huisoterapie kom voor wanneer 'n lewende wese 'n voldoende metabolisme het om 'n konstante liggaamstemperatuur te handhaaf as gevolg van die onafhanklike produksie van energie uit die verbruikte voedsel. Moderne voëls en soogdiere is ware homeotermiese wesens. Benewens voldoende energievermoë, het hulle ook verskillende meganismes wat ontwerp is om hitte te behou (vere, wol, onderhuidse laag vetweefsel) en om te beskerm teen oorverhitting by hoë omgewingstemperature (sweet). Die nadeel van hierdie meganisme is dat baie energie benodig word om die liggaamstemperatuur te handhaaf, en die behoefte aan voedsel dus hoër is as in enige ander geval.
- Traagheidsgemeenskap - dit handhaaf 'n konstante liggaamstemperatuur vanweë die groot grootte en die groot liggaamsgewig, sowel as spesifieke gedrag (byvoorbeeld in die son, koel in water). Die doeltreffendheid van die traagheid endotermie meganisme hang hoofsaaklik af van die verhouding van hittekapasiteit (vereenvoudigde - massa) en die gemiddelde hittestroom deur die liggaamsoppervlak (vereenvoudigde liggaamsoppervlakte), daarom kan hierdie meganisme slegs in groot spesies waargeneem word. Die traagheid, homo-termiese wese word stadig warm tydens periodes van temperatuurverhoging, en koel stadig af gedurende periodes van afkoeling, dit wil sê, as gevolg van die hoë hittekapasiteit, word die liggaams temperatuurskommelings uitgestryk. Die nadeel van traagheidshermoë is dat dit slegs met 'n sekere soort klimaat moontlik is - wanneer die gemiddelde omgewingstemperatuur ooreenstem met die gewenste liggaamstemperatuur en daar nie lang periodes van ernstige afkoeling of opwarming is nie. Van die voordele, moet 'n klein behoefte aan voedsel uitgelig word met 'n redelike hoë aktiwiteitsvlak. 'N Kenmerkende voorbeeld van traagheidshuisotermie is 'n krokodil. Die vel van die krokodil is bedek met 'n reghoekige, geil skilde, wat in gereelde rye op die rug en buik gerangskik is, onder in die rug en minder gereeld in die buikdeel, ontstaan osteoderms wat 'n karpataat vorm. Bedags versamel osteoderms hitte wat met sonlig binnekom. As gevolg hiervan kan die liggaamstemperatuur van 'n groot krokodil gedurende die dag binne een of twee grade wissel. Saam met krokodille kan 'n toestand naby traagheidshomeotermie waargeneem word in die grootste land- en seeskilpaaie, sowel as Komodo-akkedisse, groot pythons en boas.
Homoyotermiese diere
Homo-termiese diere (warmbloedige organismes) is diere waarvan die temperatuur min of meer konstant is en meestal nie van die omgewingstemperatuur afhanklik is nie. Dit sluit in soogdiere en voëls, waarin die konstantiteit van temperatuur geassosieer word met 'n hoër metaboliese tempo in vergelyking met poikilotermiese organismes. Daarbenewens het hulle 'n termiese isoleringslaag (verekleed, pels, vet). Hul temperatuur is relatief hoog: by soogdiere is dit 36–37 ° С, en by voëls is dit tot 40–41 ° С.
POYKILOTERM DIERE - [c. poikilos motley, diverse + termiese warmte, hitte] - koelbloedige diere, diere met 'n onstabiele liggaamstemperatuur wat wissel na gelang van die omgewingstemperatuur. Dit sluit alle ongewerweldes in, sowel as visse, amfibieë, reptiele en individuele soogdiere (vgl. homoyotermiese diere) )
Tydens evolusie het homoyotermiese diere die vermoë ontwikkel om hulself teen die koue te verdedig (migrasie, winterslaap, pels, ens.).
Ons weet reeds dat homeotermiese diere hul liggaamstemperatuur in 'n baie groter temperatuurbereik as poikilotermiese diere kan handhaaf (sien Fig. 3). Albei sterf egter by ongeveer dieselfde hoë of buitensporige lae temperature (in die eerste geval as gevolg van proteïenkoagulasie, en in die tweede - as gevolg van vries van intrasellulêre water met die vorming van yskristalle). Maar totdat dit gebeur, totdat die temperatuur kritieke waardes bereik het, sukkel die liggaam om dit op 'n normale of ten minste naby normale vlak te hou. Dit is uiteraard ten volle kenmerkend van homeotermiese organismes met termoregulering, wat die hitteproduksie en hitte-oordrag kan verbeter of verswak, afhangende van toestande. Hitte-oordrag is 'n suiwer fisiologiese proses, dit vind plaas op organiese en organismevlakke, en hitteproduksie is gebaseer op fisiologiese, chemiese en molekulêre meganismes. In die eerste plek is dit kouekoors, koue bewing, dit wil sê klein sametrekkings van skeletspiere met 'n lae doeltreffendheidskoëffisiënt en verhoogde hitteproduksie. Die liggaam skakel hierdie meganisme outomaties, refleksief aan. Die effek daarvan kan verhoog word deur aktiewe vrywillige spieraktiwiteit, wat ook hitteopwekking bevorder. Dit is geen toeval dat ons beweging gebruik om warm te bly nie.
Liggaamstemperatuur. Homotermiese diere word nie net van hitte voorsien as gevolg van hul eie hitteproduksie nie, maar kan ook hul produksie en verbruik aktief reguleer. As gevolg hiervan word hulle gekenmerk deur 'n hoë en redelik stabiele liggaamstemperatuur. By voëls is die diepste liggaamstemperatuur gewoonlik ongeveer 41 ° C met skommelinge in verskillende spesies van 38 tot 43,5 ° C (data vir 400 vvd). Onder toestande van volledige rus (hoofmetabolisme) word hierdie verskille ietwat uitgepraat, wat wissel van 39,5 tot 43,0 ° С. Op die vlak van 'n individuele organisme toon die liggaamstemperatuur 'n hoë mate van stabiliteit: die omvang van sy daaglikse veranderinge is meestal nie meer as 2-4 ° C nie, en hierdie fluktuasie hou nie verband met lugtemperatuur nie, maar weerspieël die hoeveelheid metabolisme. Selfs in Arktiese en Antarktiese spesies, by omgewingstemperatuur tot 20-50 ° C, wissel die liggaamstemperatuur binne dieselfde 2-4 ° C.
Aanpassingsprosesse by diere ten opsigte van temperatuur het gelei tot die voorkoms van poikilotermiese en homo-termiese diere. Die oorgrote meerderheid van diere is loterye, dit wil sê die temperatuur van hul eie liggame verander met die omgewingstemperatuur: amfibieë, reptiele, insekte, ens. 'N Baie kleiner deel van die diere is homo-termies, dit wil sê dat hulle 'n konstante liggaamstemperatuur het, onafhanklik van temperatuur. eksterne omgewing: soogdiere (mense ingesluit) met 'n liggaamstemperatuur van 36-37 ° С, en voëls met 'n liggaamstemperatuur van 40 ° С.
| Fisiologiese aanpassing van 'n homeotermiese dier aan koue. | 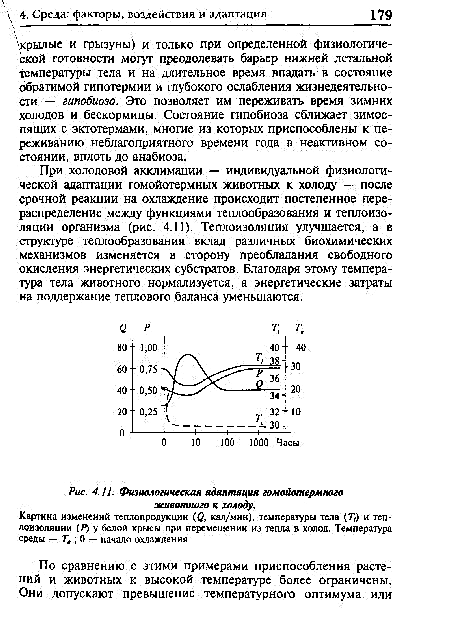 |
Maar slegs regte, warmbloedige, homeotermiese diere - voëls en soogdiere - kan 'n konstante hoë liggaamstemperatuur handhaaf met beduidende veranderinge in die omgewingstemperatuur. Hulle het perfekte senuwee- en hormonale meganismes van aktiewe hitte-regulering, wat nie net die middele vir effektiewe regulering van hitte-oordrag (deur veranderinge in perifere bloedvloei, asemhaling, sweet en hittegeleiding van die hare) insluit nie, maar ook veranderinge in die intensiteit van oksidatiewe prosesse en hitteproduksie in die liggaam. As gevolg hiervan, hang die temperatuur van die interne dele van die liggaam tot 'n beduidende mate nie van die temperatuur van die omgewing af nie. Daarom word voëls en soogdiere ook endotermiese organismes genoem. In sommige van hulle bereik termoreguleringsmeganismes groot krag. Dus verdra 'n ys, 'n uil en 'n wit gans maklik koue sonder 'n daling in liggaamstemperatuur en behou 'n verskil in die liggaam en omgewingstemperature van 100 of meer grade. Vanweë die dikte van onderhuidse vet en die eienskappe van perifere bloedsomloop, is baie pinnipeds en walvisse uitstekend aangepas vir 'n langdurige verblyf in yswater.
Aanpasbare veranderinge in hitte-oordrag by homeotermiese diere kan dus nie net daarop gemik wees om 'n hoë vlak van metabolisme te handhaaf nie, soos by die meeste voëls en soogdiere, maar ook om 'n lae metabolisme te stel in toestande wat die uitputting van energiereserwes bedreig. Hierdie vermoë om die tipes regulering van hitte-oordrag te skakel, brei die ekologiese moontlikhede op grond van homo -othermie aansienlik uit.
Aktiewe lewe by temperature onder nul kan slegs homoyotermiese diere lei. Poikilotermies, alhoewel dit temperatuur aansienlik onder nul weerstaan, maar terselfdertyd hul beweeglikheid verloor. Die meeste diere is 'n temperatuur van +40 ° C, dit wil sê selfs laer as die koagulasietemperatuur van die proteïen.
Tydens Koue Auslimation - individuele fisiologiese aanpassing van homeotermiese diere aan koue - vind daar na 'n dringende reaksie op verkoeling 'n geleidelike herverdeling plaas tussen die funksies van hitteopwekking en termiese isolasie van die liggaam (Fig. 4.11). Termiese isolasie verbeter, en in die struktuur van hitteopwekking, verander die bydrae van verskillende biochemiese meganismes in die rigting van die voorkoms van vrye oksidasie van energiesubstrate. As gevolg hiervan word die liggaamstemperatuur van die dier genormaliseer, en word die energiekoste verbonde aan die handhawing van die hittebalans verminder.
'N Fundamenteel verskillende tipe aanpassing by die temperatuurfaktor is kenmerkend van homo-termiese diere. Hul temperatuuraanpassings hou verband met die aktiewe instandhouding van 'n konstante interne temperatuur en is gebaseer op 'n hoë vlak van metabolisme en 'n effektiewe regulerende funksie van die sentrale senuweestelsel. Die kompleks van morfofisiologiese meganismes om die liggaam se termiese homeostase te handhaaf, is 'n spesifieke eienskap van homeotermiese diere.
As poikilotermies gevoelloos is, dan is die winter- en somerwinterslaap inherent aan homo-termiese diere, waarvan die fisiologiese en molekulêre meganismes verskil van gevoelloosheid. Hul eksterne manifestasies is dieselfde: 'n afname in liggaamstemperatuur byna tot omgewingstemperatuur (slegs gedurende die winterwinterslaap, gedurende die somerwinterslaap is dit nie) en die metaboliese tempo (10-15 keer), 'n verskuiwing in die reaksie van die interne omgewing van die liggaam op die alkaliese sy, 'n afname in die opgewondenheid van die asemhalingsentrum en afneem in asemhaling tot 1 inspirasie binne 2,5 minute, die hartklop daal ook skerp (byvoorbeeld in vlermuise van 420 tot 16 slae / min). Die rede hiervoor is 'n toename in die toon van die parasimpatiese senuweestelsel en 'n afname in die opgewondenheid van die simpatiese. Die belangrikste is dat die termoreguleringstelsel tydens die winterslaap afgeskakel is. Die redes hiervoor is 'n afname in die aktiwiteit van die skildklier en 'n afname in die inhoud van skildklierhormone in die bloed. Homoyothermic diere word poikilothermic.
Voëls en soogdiere kan 'n redelike konstante liggaamstemperatuur handhaaf, ongeag die omgewingstemperatuur. Hierdie diere word homokotermies genoem (van Grieks. Homoyotermiese diere is relatief min afhanklik van eksterne hittebronne. As gevolg van die hoë wisselkoers, genereer hulle 'n voldoende hoeveelheid hitte wat geberg kan word. Aangesien hierdie diere bestaan as gevolg van interne hittebronne, word hulle dikwels endotermies genoem) .
Al die bogenoemde verwys na die sogenaamde diep liggaamstemperatuur, wat die termiese toestand van die termostatiese beheerde “kern” van die liggaam kenmerk. By alle homo-termiese diere vorm die buitenste lae van die liggaam (integument, deel van die spiere, ens.) 'N min of meer uitgesproke “dop” waarvan die temperatuur baie verskil. Dus kenmerk 'n stabiele temperatuur slegs die gebied van lokalisering van belangrike interne organe en prosesse. Oppervlakweefsel is meer bestand teen temperatuurskommelings.Die ego kan nuttig wees vir die liggaam, aangesien die temperatuurgradiënt by die grens van die liggaam en die omgewing in so 'n situasie afneem, wat dit moontlik maak om termiese homeostase van die "kern" van die liggaam te handhaaf met laer energie-uitgawes.
Die vrystelling van energie in die vorm van hitte gaan gepaard met die funksionele las van alle organe en weefsels (Tabel 4.2) en is kenmerkend van alle lewende organismes. Die spesifisiteit van homeotermiese diere is dat 'n verandering in hitteproduksie as reaksie op 'n veranderende temperatuur 'n spesiale reaksie van die liggaam is wat nie die funksioneringsvlak van die basiese fisiologiese stelsels beïnvloed nie.
LANDSKAP HOMEOSTASIS Die vermoë van 'n landskap om sy struktuur en die aard van die verbindings tussen elemente te behou ten spyte van eksterne invloede, is basies. HUIS-TERMALE DIERE [vanaf c. Iotoyuz is soortgelyk, identies en (Yeghts - hitte), warmbloedige diere - diere wie se liggaamstemperatuur konstant gehou word, ongeag die omgewingstemperatuur as gevolg van die energie wat tydens die metabolisme vrygestel word (voëls en soogdiere).
Die effek van die omgewingstemperatuur. Die konstansie van liggaamstemperatuur, (homo-termiese) diere, is noodsaaklik vir die ontwikkeling en lewensbelangrike aktiwiteit van weefsels, organe en die liggaam as geheel. Homo-termiese diere word onderskei deur die evolusionêr ontwikkelde vermoë om die hoeveelheid hitte-oordrag (fisiese termoregulering) te verander deur die bloedsomloop in die oppervlakweefsel te reguleer en die vog van die liggaam te verdamp, asook om hitteopwekking (chemiese termoregulering) te verander terwyl 'n konstante temperatuur van die weefsels en die hele liggaam gehandhaaf word. Die relatiewe konstansie van die liggaamstemperatuur van mak diere word ondersteun deur ingewikkelde, neurohumorale regulering van die prosesse van hitteopwekking en hitte-oordrag. Wanneer die liggaam in die liggaam afkoel, neem metaboliese prosesse toe en word hitteopwekking toeneem, en hitte-oordrag neem af, as dit verhit word, inteendeel, die hitteproduksie neem af, en die hitte-oordrag neem toe.
Spesifieke verskille in die temperatuurdrempel waaronder die normale funksionering van die spermbewegingsapparaat versteur word, veral as die sperm van poikilotermiese en homo-termiese diere vergelyk word, kan op verskillende maniere verklaar word (Holwill, 1969). Eerstens kan verskillende organismes variasies hê in die struktuur van die ensiem, die aantal en tipe bindings wat beskadig word deur termiese denaturering van die molekules. Tweedens kan die ensiem in die bestudeerde diersoort identies wees, en verskille in temperatuurgrense waarteen die denaturering daarvan waargeneem word, is waarskynlik te wyte aan die uiteenlopendheid van omgewingstoestande (pH, ioonkonsentrasie, ens.).
Lug as 'n leefomgewing het sekere kenmerke: wat die algemene evolusionêre paaie van die inwoners van hierdie omgewing lei. Dus bepaal 'n hoë suurstofinhoud (ongeveer 21% in die lug in die lug, effens minder in die lug wat die asemhalingstelsel van diere vul) die moontlikheid om 'n hoë vlak van energiemetabolisme te vorm. Dit is geen toeval dat daar in hierdie omgewing homo-termiese diere na vore gekom het nie, gekenmerk deur 'n hoë vlak van energie in die liggaam, 'n hoë mate van outonomie van buite-invloede en 'n hoë biologiese aktiwiteit in ekosisteme. Aan die ander kant word atmosferiese lug gekenmerk deur lae en wisselende humiditeit. Hierdie omstandigheid het die moontlikhede vir die ontwikkeling van die lugomgewing grootliks beperk, en onder die inwoners is dit gelei deur die evolusie van die fundamentele eienskappe van die watersoutmetabolisme en die struktuur van die asemhalingstelsel.
Die tweede belangrike omgewingsvoordeel vir die inwoners van lewende organismes is hul beskerming teen die direkte impak van omgewingsfaktore. Binne-in die gasheer het hulle feitlik geen gevaar vir uitdroging nie, skerp temperatuurskommelings, wesenlike veranderinge in sout en osmotiese regimes, ens. Daar is dus binne veral stabiele toestande interne inwoners van homo-termiese diere. Skommelinge in omgewingstoestande beïnvloed interne parasiete en simbionte slegs indirek deur die gasheerorganisme.
Die mens as 'n spesie, fundamenteel verskillend van alle vorige spesies, het ontstaan tydens die evolusieproses onder die invloed van wette wat gemeenskaplik was aan alle lewende dinge as gevolg van 'n fundamentele geneties-vaste ontdekking in die evolusieproses van die organismes van die biosfeer. Sulke kardinale ontdekkings, wat gelei het tot die opkoms van fundamenteel nuwe spesies, het voor die mens verskyn. Daar was dus meersellige organismes, gewerweldes, homeotermiese diere met 'n konstante liggaamstemperatuur.
Die voorbeelde wat hierbo gelys is, is ver van alle vorme van aanpasbare gedrag. Dit sal die vermoë van baie voëls en soogdiere toevoeg om aktief neste, gate en ander skuilings te bou met 'n gunstige mikroklimaat, die gebruik van houers wat energieverbruik bespaar, seisoenale bewegings, die aanpasbare aard van daaglikse aktiwiteite, ens. Die hele kompleks van aanpasbare gedragsreaksies, wat die spanning van energie-uitruiling verminder, brei die ekologiese vermoëns van homeotermiese diere uit.
Die geassimileerde energie minus die energie wat in die geskei is wat uit die liggaam geskei word (ontlasting, urine, ens.) Is die gemetaboliseerde energie. 'N Deel daarvan word in die vorm van tesha toegewys in die verwerking van voedsel en word versprei óf gebruik vir termoregulering. Die oorblywende energie word verdeel in die bestaansenergie, wat onmiddellik deur die algemeenste lewensvorme verbruik word (in wese is dit ook 'uitgawes aan asemhaling'), en die produktiewe energie, wat (ten minste tydelik) opgehoop word in die massa groeiende weefsels, energiereserwes en seksuele produkte (rys 3.1). Die energie van bestaan bestaan uit die koste van fundamentele lewensprosesse (basale metabolisme, of basale metabolisme) en die energie wat aan verskillende vorme van aktiwiteit bestee word. By homo-termiese diere word die energie-uitgawes aan termoregulering hieraan toegevoeg. Al hierdie energiekoste eindig met die verspreiding van energie in die vorm van hitte - weereens omdat daar nie 'n enkele funksie met 'n doeltreffendheid van 100% werk nie. Die opgehoopte energie in die weefsel van die heterotrofe liggaam vorm die sekondêre produksie van die ekosisteem, wat verbruikers van hoër bestellings as voedsel kan gebruik.
Die voordele van homeotermie
Warmbloedige diere val in die reël nie in 'n winterslaap nie, behalwe op enkele uitsonderings, en hulle kan dwarsdeur die jaar aktief wees, hulself eet, beweeg en beskerm teen roofdiere.
Alhoewel warmbloedige diere baie voedsel moet verbruik om aktief te bly, het hulle die energie en die middele om alle natuurlike gebiede te oorheers, selfs in koue Antarktika of hoë bergreekse. Hulle kan ook vinniger en oor langer afstande reis as koelbloedige diere.
Nadele van homeotermie
Aangesien die liggaamstemperatuur by warmbloedige diere stabiel bly, is dit die ideale gasheer vir baie parasiete, soos wurms, of mikro-organismes, insluitend bakterieë en virusse, waarvan baie dodelike siektes kan veroorsaak.
Aangesien homotermiese diere hul eie hitte vrystel, is 'n belangrike faktor die verhouding van massa tot liggaamsoppervlakte. 'N Groter liggaamsmassa produseer meer hitte, en 'n groter liggaamsoppervlak word in die somer of in 'n warmer habitat, soos die groot ore van olifante, afgekoel. Daarom kan warmbloedige diere nie so klein soos koudbloedige insekte wees nie.